|
随着社会老龄化的加速,心脑血管病已成为严重威胁人类健康的重大疾病,其最主要的病变基础是动脉粥样硬化(atherosclerosis, AS)。血管平滑肌细胞(vascularsmooth muscle cell, VSMC)涉及到AS病变发生、发展的全部过程。VSMC由收缩表型转化为合成表型的过程称之为表型转化。VSMC表型转化是易损斑块、支架术后再狭窄和动脉粥样硬化等血管性病变发生发展过程中的关键性步骤[1]。 巨细胞病毒(cytomegalovirus,CMV)属于疱疹病毒科β亚科,是一种广泛存在于自然界的机会致病性病毒。既往研究证实CMV所致的慢性潜伏性感染与动脉粥样硬化的形成发展密切相关[2]。原发性CMV感染首先累及内皮细胞,随后在VSMC内潜伏或持续性感染,导致VSMC的肥大、增殖和迁移,对AS的形成及狭窄起重要作用[3],但CMV感染是否参与VSMC表型转化的过程目前尚未明确。 PI3K/Akt信号通路是调节VSMC表型转化的主要信号通路[4],有文献报道,其下游转录激活因子FoxO通过与促血管平滑肌细胞分化因子Myocardin的协同作用来调控表型转化标志基因的表达。 本实验以平滑肌22α(smooth muscle 22 alpha,SM22α)和骨桥蛋白(osteopontin,OPN)分别作为VSMC收缩型和合成型的标志基因,旨在通过体内和体外实验证实CMV是否参与VSMC的表型转化过程,以及其可能的作用途径和机制。
1.1 制备小鼠巨细胞病毒(murine cytomegalovirus,MCMV) 感染致动脉粥样硬化的apoE-/-小鼠模型MCMV Smith株购自美国典型生物收藏中心(American Type Culture Collection,ATCC),通过空斑形成实验确定小鼠巨细胞的滴度为2×105 PFU。8周龄雄性apoE-/-小鼠,体质量(20±2)g,购自北京维通利华实验动物技术有限公司,实验动物生产许可证号SCXK(京)2014-0004,SPF级环境,高脂饲料喂食(购自北京科澳饲料有限公司,21%猪油,1.2%胆固醇,0.2%胆酸钠)。适应性饲养1周后开始实验。将动物随机分为感染组和正常对照组,每组12只,感染组予2×105 PFU MCMV 200μl腹腔注射,对照组予等量的DMEM腹腔注射。分别于实验第8、12、16周从两组各随机选取1只小鼠,通过超高分辨率小动物彩色多普勒超声成像系统(MS-550高频宽带电子线阵探头,频率40 MHz)判断小鼠主动脉粥样硬化斑块形成情况,决定取材时间。 1.2 apoE-/-小鼠主动脉取材、HE染色及免疫组化检测 ApoE-/-小鼠禁食水8 h后称重,取血。麻醉后取腹侧正中线切口,暴露主动脉及其分支。切取主动脉起始至左锁骨下动脉开口段,4%多聚甲醛固定,石蜡包埋,5 μm连续切片。Image-Pro Plus 6.0图像分析软件采集apoE-/-小鼠主动脉动脉粥样硬化斑块面积、管腔面积。每个指标选取4个不同切面,取平均值进行统计。免疫组化检测各组小鼠主动脉斑块中平滑肌表型转化的标志蛋白SM22a和OPN的表达。 1.3 小鼠主动脉血管平滑肌细胞(MOVAS)的培养 MOVAS细胞购自美国ATCC,用含10%PBS的细胞培养液,置于37 ℃,5%CO2细胞培养箱中培养,待细胞融合至70%~80%,用0.25%胰酶和0.02%EDTA消化,按1∶4传代,选取第3~5代进行实验。MOVAS细胞分为两组,实验组将MCMV与MOVAS共同孵育,对照组加入等量的DMEM,24 h后收集细胞进行实验。 1.4 小鼠主动脉血管平滑肌细胞(MOVAS)表型转化相关指标的检测 应用不同滴度MCMV刺激MOVAS细胞,MTT法检测不同时间点各滴度MCMV对MOVAS细胞增殖的影响。选取MOVAS细胞增殖最明显的MCMV滴度作为实验组,并以正常MOVAS细胞作为对照组。应用Western blot检测两组细胞表型转化标志蛋白SM22a和OPN的表达。 1.5 MCMV对MOVAS表型转化PI3K/Akt通路的作用 应用Western blot分别检测对照组和实验组PI3K/Akt通路中的Akt、磷酸化的Akt表达水平。给予特异性的PI3K通路的抑制剂Ly294002后,应用Western blot检测两组平滑肌细胞表型转化标志蛋白SM22a和OPN的表达情况。 1.6 统计学方法 采用SPSS 16.0软件进行统计学分析。计量数据以±s表示,组间比较采用t检验。P<0.05表示差异有统计学意义。
2.1 apoE-/-小鼠主动脉HE染色及免疫组化结果 实验第16周,小动物超声检查显示感染组主动脉壁内膜增厚,连续性中断,并可见动脉粥样硬化斑块形成,斑块内回声略高于内膜。对照组未见明显斑块形成。因此,本研究选择饲养16周时进行主动脉取材。apoE-/-小鼠主动脉HE染色观察脂质斑块发现,对照组和感染组小鼠16周时动脉斑块面积分别为(3.16±0.72)%和(28.71±3.07)%,感染组的动脉粥样硬化斑块的程度较正常饮食组严重(P=0.007,图1),且二者相比差异有统计学意义。
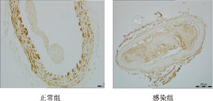
图1 饲养16周时正常组和感染组apoE-/-小鼠主动脉HE染色(×40) Fig.1 HE staining of aorta in control andinfection group of apoE-/- mice after 16 weeks of feeding (×40) 免疫组化染色结果显示,对照组的10个主动脉中有9个SM22a染色阳性,SM22a阳性染色颗粒主要位于血管壁富含平滑肌细胞的中膜和外膜,而OPN染色阳性的主动脉则只有1个。CMV感染组的结果与对照组相反,10个主动脉中2个SM22a染色阳性,而OPN染色阳性的主动脉则多达10个,OPN阳性染色颗粒主要位于血管壁的中膜、外膜以及粥样硬化斑块中(图2、图3)。在未出现明显动脉粥样硬化斑块的对照组中,动脉壁以表达收缩期蛋白为主;在MCMV感染组,动脉粥样硬化程度加重,动脉壁中高表达的收缩期蛋白被合成期的蛋白所取代,说明MCMV感染促使动脉粥样硬化进展的同时,诱导了平滑肌细胞的表型转化。
图2 正常组(×100)和感染组apoE-/-小鼠主动脉SM22a免疫组化染色(×40) Fig.2 SM22a immunohistochemical staining ofaorta in normal group (×100) and infection group of apoE-/-mice(×40)
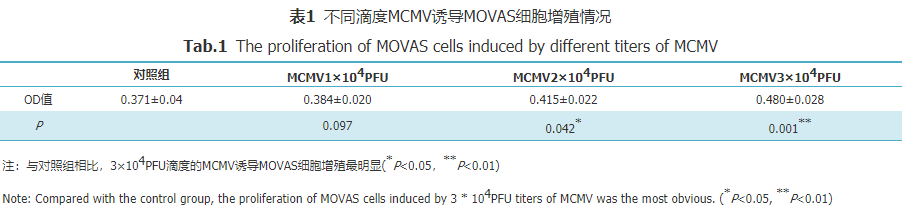
图3 正常组和感染组apoE-/-小鼠主动脉OPN免疫组化染色(×100) Fig.3 OPN immunohistochemical staining ofaorta in normal group and infection group of apoE-/- mice(×100) 2.2 MCMV感染对MOVAS表型转化指标的影响 观察不同滴度及时间下MCMV对MOVAS细胞增殖能力的影响,发现在24 h诱导3×104PFU滴度的MCMV诱导细胞增殖最明显(表1)。
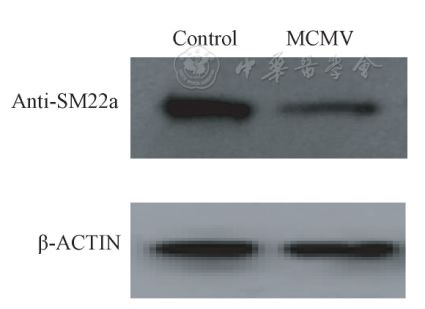
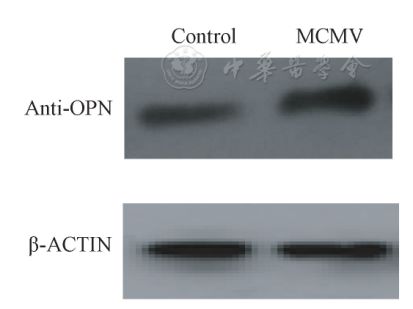
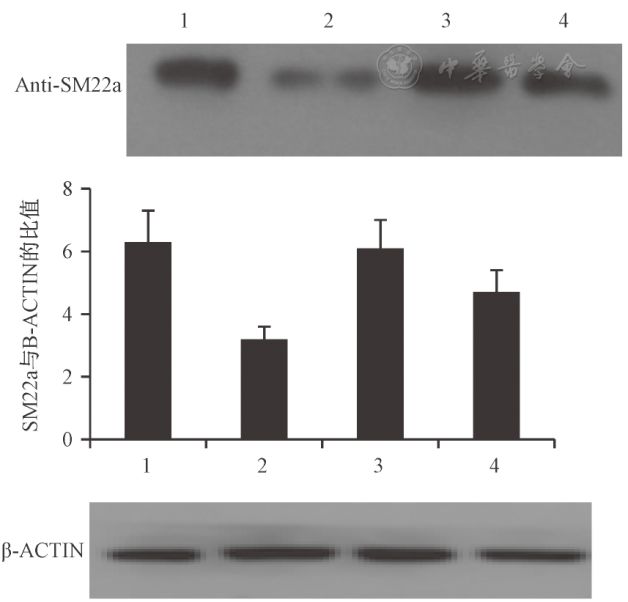
表1不同滴度MCMV诱导MOVAS细胞增殖情况 Tab.1 The proliferation of MOVAS cellsinduced by different titers of MCMV Western blot结果显示,3×104PFU MCMV感染MOVAS24 h细胞后,SM22a表达较对照组下降,OPN表达较对照组增加,且两组与对照组相比差异均有统计学意义(P=0.023,P=0.034,图4、图5)。
图4 SM22a蛋白在对照组和实验组的表达 Fig.4 Expression of SM22a protein in thecontrol group and the experimental group
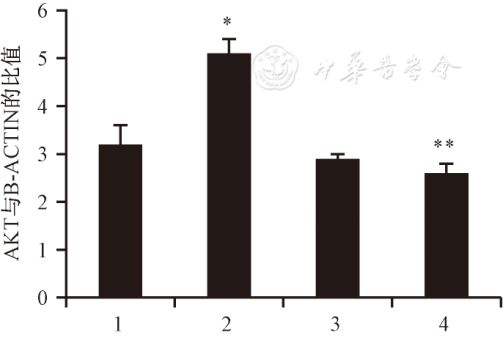
图5 OPN蛋白在对照组和实验组的表达 Fig.5 Expression of OPN protein in controlgroup and experimental group 2.3 MCMV对MOVAS表型转化PI3K/Akt通路的调控 为明确PI3K/Akt通路在MCMV促平滑肌细胞表型转化中的作用,本研究检测了磷酸化的Akt蛋白的表达水平,结果发现MCMV明显刺激磷酸化Akt蛋白表达上调(P=0.035)。本研究预先用PI3K的抑制剂Ly294002(10-6mol/L)孵育MOVAS30 min后,用3×104PFU滴度的MCMV刺激24 h, LY294002可阻断MCMV感染后的Akt蛋白磷酸化(P=0.031),而对对照组无明显影响(P=0.081,图6),提示MCMV促平滑肌细胞表型转化可能经由PI3K/Akt信号通路介导。
图6 LY294002预处理前后,对照组和感染组磷酸化的Akt蛋白表达 Fig.6 Expression of phosphorylated Akt proteinin control group and infection group before and after LY294002 pretreatment 图7显示单独LY294002对MOVAS表型转化收缩期标志蛋白SM22a的表达无影响,但MCMV+LY294002组的SM22a蛋白表达较MCMV组明显上调,且与对照组相比统计学差异消失。说明LY294002可明显抑制MCMV促平滑肌细胞表型转化的作用,进一步提示PI3K/Akt信号途径可能参与了MCMV促平滑肌细胞表型转化的过程。
图7 LY294002预处理前后,对照组和感染组MOVAS细胞中SM22a蛋白的表达 Fig.7 Expression of SM22a protein in MOVAScells of control group and infection group before and after LY294002pretreatment
VSMCs的表型决定了细胞的生物学特性和功能[5]。正常VSMCs是收缩型,VSMCs表型由收缩型向合成型转化是其增生的先决条件。生理情况下,VSMC处于非增殖状态,表现为分化良好的收缩表型。血管损伤后,VSMC转化为增殖性的合成表型,合成和分泌多种血管活性物质;同时自身发生肥大,增殖和迁移,导致管壁增厚、管腔狭窄。SM22α是一个22×103的细胞骨架蛋白,其功能涉及血管平滑肌细胞的收缩调节,被认为是收缩表型的标志物之一。骨桥蛋白(osteopontin,OPN)是细胞外基质中一种富含唾液酸的磷酸化糖蛋白,最早从骨基质分离,是合成型细胞的标记基因,它的出现代表血管平滑肌细胞进入了合成型,具有了活跃的增殖与迁移能力。电镜下,典型的收缩表型和合成表型的VSMCs应具有明显的形态、结构区别。前者核小、染色质致密,胞浆富含收缩蛋白、肌丝,而较少内质网、高尔基复合体等合成、分泌性细胞器。而后者则核大、染色质疏松,少肌丝,但却富含合成、分泌性细胞器。 CMV感染首先累及内皮细胞,随后在VSMC内潜伏或持续性感染,导致VSMC的肥大、增殖和迁移[6]。研究发现,在CMV所致的apoE-/-小鼠动脉粥样硬化模型中,收缩表型标志蛋白SM22α较对照组表达明显减少,而合成表型标志蛋白OPN的表达则明显增加。细胞学实验结果与之相符,说明CMV感染可以促进平滑肌细胞发生表型转化。 VSMC表型转化受多重因素的调控,PI3K/Akt信号通路是调节VSMC表型转化的主要信号通路。在本实验中,PI3K/Akt信号通路的阻断剂LY294002可以显著阻断MCMV感染诱导的SM22a蛋白表达的下调,以及Akt蛋白的磷酸化,而对照组无明显变化。有文献报道,PI3K/Akt信号通路的下游产物FoxO通过与促VSMC表型转化的转录激活因子Myocardin的协同作用来调控表型转化标志基因的表达[7]。之前的研究发现CMV通过编码miR-217下调FoxO3a蛋白的表达,使血管内皮细胞增殖、迁移能力明显增强[8]。近期的研究还发现,Myocardin的转录活性能被CMV所增强,促使VSMC发生表型转化[9],这也为进一步探讨CMV诱导的平滑肌细胞表型转化的作用机制提供了思路。 总之,CMV感染可以促使平滑肌细胞发生表型转化,PI3K/Akt信号通路是其作用途径,但其详细的作用机制,目前还远未明确。 |
Copyright © 2015-2023 杭州宇翼科技有限公司 丨 Discuz! X3.5 丨增值电信业务经营许可证:浙B2-20190572丨浙ICP备18026348号-1丨浙公网安备33010802009352号